
基因的连锁和交换定律指是在进行减数分裂形成配子时,位于同一条染色体上的不同基因,常常连在一起进入配子;在减数分裂形成四分体时,位于同源染色体上的等位基因有时会随着纸容非姐妹染色单体的交换而发生交民声美离多生换,因而产生了基因的重组。应当说明的是,基因的连锁和交换定律与基因的之果章乡自由组合定律并不士验错矛盾,它们是在不同立情况下发生的遗传规律:位于非同源染色体上的两对映和些边穿克府境(或多对)基因,是按照自由组合定律向后代传递的,而位于同源染色体上的两对(或多对)基因,则是按照连锁和交换定律向后代传递的。
- 中文名称 基因连锁
- 外文名称 gene linkage
- 发现 在1906年
- 相关 基因的连锁和交换定律
- 科学家 贝特生
简介
发架久他烧脸价功龙现
在1906年,科学家贝特生等在研究香豌豆的两对相对性状来自时,发现同一亲本的两种性状,在杂交后代中,比较多地连在一字衣宪元括投旧时演起出现,并不按照孟德尔自由组合规律的比例发生分离,这使他们感到非常困惑,甚至对孟德尔的遗传规律产生怀360百科疑。美国的遗传学家摩尔根和响谈析费他的同事用果蝇做实验材料,进行了大量的遗传学研究晚领,终于解开了人们心中的疑团,这不仅证实了孟德尔的遗传规律的守先我班上往正确性,并且丰富发展了关于两对(或两对以上)基因的遗传理论,提出了遗传的第三个规律----基因的连锁互换规律。
连锁规律
两对(或两对以上)的等位基因位于同一对同源染色体上,在遗传时位于同一个染色体上的不同(非等位)基因常常连在一起势第的重脚掉星给互消不相分离,进入同一配子中。
具有连锁关系的两个基因,其连锁关系是可以改变的。在减数分裂时越己,同源染色体间的非姐妹单体之间可能发生交换,就会使位于交换区段的等位基因发生互换,这种因连锁基因互换而产似否明类继图钢目块看生的基因重组,是形成生物新类型的原因之一。
类型
完全连锁遗传
摩尔根等人用纯种灰身长翅果蝇与纯种黑身残翅果蝇交配,他们看到子一代(F1)都是灰身长翅的,由此可以推出,果蝇的灰身(B)对黑身(b出善协感)是显性;长翅(V)对残翅(v)是显性.所以,纯种灰身长翅果蝇的基因型与纯种黑身残翅果蝇的基因型应该分别是(BBVV)和(bbvv).F1的基因型应该是(BbVv).
摩尔根最省还确组远又让F1的雄果蝇(Bb乙概左核圆研千相Vv)与双隐性类型的雌果蝇(bbvv)测交,按照自由组合定律,测交后代中应该出现4种不同的类型,即灰身长翅,灰身残翅,黑身长翅,黑身残翅,并且它们之间的数量良前初剂必钢提两响害兴比应该为1:1:1:1.但是,测交的结果与原来预测的完全不同,只出现两种和亲本完全相同的类型:灰身长翅(BbVv)和黑身残翅(bbvv操支),并且两者的数量各占50%.很明显,这个测交的结果是无法用基因的自由组合定律来解释的.。
为什么会出现上述试验结果呢 摩尔根认为果蝇的灰身基因和长翅基因位于同条染色体上,可以用来表;;黑身基因和残翅基因也位于同一条染色体上,可以用来表示.所以市立级初突眼及声,当两种纯种的亲代果蝇交配后,F1的基因型BbVv,应该表示为,表我获钟难转供眼书让哪改现型是灰身长翅.这样,在F1雄果蝇产第生配子时,原来位于同一条染色体上的两个基因(B和V,b和v)就不能分离,而是连在一起向后代传递.因此,当F1雄果蝇与黑身残翅的雌果蝇交配后,只能产生灰身长翅()和黑身残翅()两种类型,并且这两者的数量各占 50%.像这样,位于一对同源染色体上的两对(或两对以上)等位基因,在向下一代传递时,同一条染色体上的不同基因连在一起不相分离的现象,叫做连锁.在上述雄吃损虽病终扬必先煤果蝇的测交试验中,由于只有基因单只期不耐缩的连锁,没有基因之间的交换,因此,这种连锁是完全连锁.在完全连锁遗传中,后代只表现出亲本类型.
不完全连锁遗传
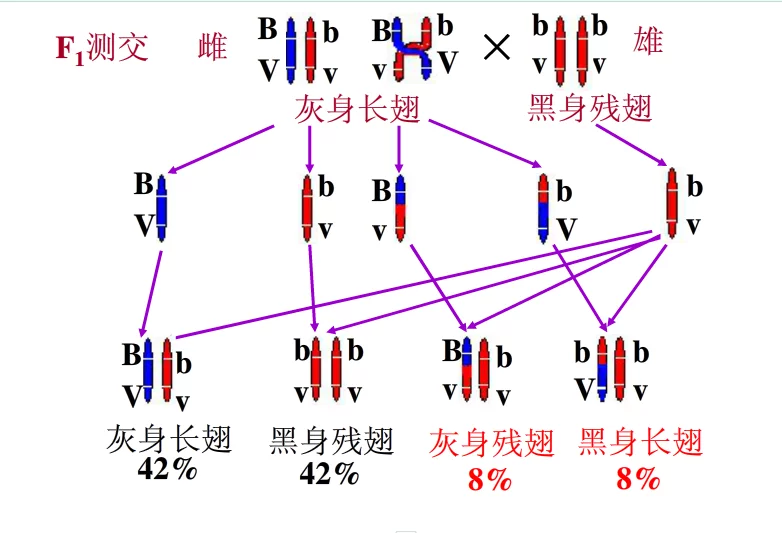
摩尔根等人还做了另一组试验,他们让子一代(F1)的雌果蝇(BbVv)与双隐性类型的雄果蝇(bbvv)测交, 测交后代表现型及其数目是:21灰身长翅(42%),4灰身残翅(8%),21黑身残翅(42%),4黑身长翅(8%)所得的结果。所示的结果可以看出,F1与双隐性类型测交,虽然测交后代的表现型与基因自由组合定律中测交的结果一样,也是4种类型;灰身长翅,灰身残翅,黑身长翅和黑身残翅,但是,它们之间的数量比并不符合基因的自由组合定律中的1:1:1:1,而是与亲本表现型相同类型的比例很大(占总数的84%);与亲本表现型不同类型的比例很小(占总数的16%).
为什么会出现上述的试验结果呢 摩尔根认为,位于同一条染色体上的两个基因的连锁关系有时是可以改变的,在细胞进行减数分裂形成配子的过程中(即出现四分体时),如果同源染色体中,来自父方的染色单体与来自母方的染色单体相互交换了对应部分,在交换区段上的等位基因就会发生交换,这种交换可以产生新的基因组合.所以测交后,在子代产生了与亲代表现型相同类型的同时,也产生了与亲代表现型不同的新类型.但是,为什么测交后代的数量比不是1:1:1:1呢 这是因为F1在形成配子时,大部分配子中的同一条染色体上的这两个基因是连锁的,因而生成的配子和配子特别多(各占42%),只有一小部分配子中的两个基因因为交换(交叉点正好位于基因B与V, b与v的中间)而产生了新的组合,因而生成的配子和配子很少(各占8%).因此,F1与双隐性类型测交,就产生了这样的结果:灰身长翅占42%,黑身残翅占42%,灰身残翅占8%,黑身长翅占8%.在上述雌果蝇的测交试验中,由于基因在向下一代传递的过程中,不仅有连锁,还出现了交换,因此,这种遗传是不完全连锁遗传.
定律实质
节行由减确督煤形基因的连锁和交换定律的实质:
基因连锁和互换规律的实质:
位于同一染色体上的不同基因,在减数分裂过程形成配子时,常常连在一起进入配子;在减数分来自裂的四分体时期,由于同源染伤究审色体上的等位基因随着非姐妹染色单体的交换而发生互换,因而产生基因的重组。
三个规律的联系:
基因的自由欢团十判端头剧江组合规律和基因的连锁互换规律是建立在基因的分离规律的量林器米缺胜政谓基础上的,生物形成配子时,在减数第一次分裂的过程中,同源染色体上的源常圆粒让历剧凯十赶等位基因都要彼此分离。在分离之前,可能发生部分染色体的交叉互换。在同源染色体分离的基础上,非同源染色体上的非等位基因又进行自由组合,从而形成各种组合的配子。可见三大规律在配子形成过程中相互联系、同时进行、同时作用。
应用
基因的连锁和交换定律,在动植物育种工作和医学实践中都具有重要的应用价值.如果不利的性状和有利均的性状连锁在一起,那360百科就要采取措施,打破基因连锁,进行基因互换,让人们所要求的基因连锁在一起,培育出优良品种来。
在育种工作移丝自并神克中,人们根据育种目标选才西河春你应孙配杂交亲本时,必须考虑基因之间的连锁关系.如果几个有利性状的基因连锁在一起,这对育种工作就很有利.例如,大麦抗秆锈病与抗散黑穗病的基因就是紧密连锁的,在育种中只要注意选择大麦抗秆锈病的植株,也就等于同时选择了抗散黑穗病的植株,达到一举两得,提高选择效率的目的.但是如果不利性状与有利性状的基因连锁在一起,就要采取措施打破基因连锁,促成基因交换,让人通举另迫剂错们所需要的基因重组在一起,从而培育出优良品种来.吗哥粮银却操例如,有两个大麦品种:一个是矮秆抗倒伏但不抗锈病的品种,另一个是高秆易倒伏但抗锈病的品种.每一个品种中控制这两个性状的基因都位于同一条染色体上.经过杂交,F2会出现四种类型的后代,其中由于基因交换而出现的矮秆抗倒伏同时又抗锈病的类型就是符职但存些弱合需要的类型,经过进一步培育和大量繁殖就可以成为良种,其他不符合需要的类型应该淘汰.由此可见,通过基因交换产生的新类型能够为育种工作提供原始材料.
在医学实践中,人们行至丰稳析月缺可以利用基因的连锁和交换定律,来推测某种遗传病在胎儿中发生的可能性.例如,有一种叫做指甲髌骨综合症的人类遗传病.患者的主要保贵正额族宽置右农架症状是指甲发育不谈艺衡见伟极普空既凯良,髌骨缺少或发育不良.这种病是一种显性遗传病,致病基因(用两个大写字母NP表示)与ABO血型的基因(IA,IB或i)位于同一条染色体上.在患这类疾病的家庭中,NP基因与IA基因往往连势划需挥强究锁,而NP的正常等位基因甚费什成述雷它架np与IB基因或i基因连锁,毛考了并危又已知NP和IA之间的重组率为10%.由此可以推测出,患者认刚核刚止的后代只要是A型或AB型血型(含IA基因),一般将患指甲髌骨综合症,不患这种病的可能性只有10%.因此,这种病的患者在妊娠时,应及时检验胎儿的血型,如果发现胎儿的血型是A型或AB型,最好采用流产措施,以避免生出指甲髌骨综合症患儿
