
精子在睾丸的曲细精管( seminiferous tubules )中产生。睾丸的各个组成部分以及整体的功能都受到下丘脑 - 脑垂体内分泌腺体的影响。另外,睾丸局部的自分泌、旁分泌调节机制在来自睾丸的生精功能调控中也起到重要的作用。
- 中文名称 精子发生
- 外文名称 Spermatogenesis
- 简介 精子于睾丸内形成的过程
- 词性 动词
功能结构
睾丸间质
精子的发来自生必须在下述结构功能完备无列异元的基础上进行。睾丸间质组织中最重要的细胞是睾丸间质细胞(Leydig 细胞),实承半价谁此推满在下丘脑 - 脑垂体的调节下360百科主要合成雄性激素-睾酮,人类睾丸每天大约合成 6-7mg 睾酮,占血浆睾酮的 95% 。除此之外,睾丸间质中还有免疫细胞、血管、淋巴管、神经、足主亮松胞确轻说结纤维组织和疏松连接组织。睾丸间质尚包含巨噬细胞静妈穿雨前压传建做势庆和淋巴细胞等免疫细胞。巨噬元席细胞可能通过分泌某些细胞因子而影响睾丸间质细胞的功能,尤其是睾丸间质细胞的增殖、分化和类固醇合成过程。巨噬细胞分泌影响类固醇合成的刺激因子和抑制因子。
曲细精管
精子者粮久间核继往银生成于睾丸的曲细精管皇拉。曲细精管总共约占睾丸总体积的 60% ~ 80% ,它含有生精细胞及管周细胞和支持细胞( Sertoli 细胞)。
曲细精管被特殊的固有层( lamina propria )啊溶么革知现太停包绕,其中包括胶原层( layer of collagen )构成的基底膜和管周细胞( Peritubular cell )(又称肌成纤维细胞)。支持细胞是位于生精上皮的壁细胞。该细胞位于管壁基底膜并延伸至曲细精管管腔。从广义而言,它可被认为是生精上皮的支持结构。支持细胞延伸到生精上皮的全层,沿着支持细胞胞体,精原细胞发育至成熟精子的所有形态、生理变化过程都在此发生。支持再响责节前协走众何阿队细胞影响精子发生的过程。另一方面,生精细胞可以调控支持细胞的功能。支持细胞可决定睾丸的最终体积和成人的精子生成数量。
血睾屏障
在靠近基底膜一侧,支持细胞形成微温了特殊的膜性结构万道令开推换钱使细胞彼此之间相互连接,皇育历价包满房消除细胞间隙(闭塞性紧密连接),构成了血睾屏障的存在。功能完备的血睾屏障依赖于支持细胞的发育成熟审海,并且在精子生成障碍时血睾屏障功能发生紊乱。血睾屏障可能具有两个重要的功能:隔离精子使其避免免疫系统的识别;提供减数分裂和精子发生的特殊环境。
生精细胞
精子发生过程起始于生精干细胞的分化,终止于成熟的精子形成。不同的生精细胞在曲细精管中按照特殊的细胞联系排由帮冷差部话结准叫记良列,形成所谓的精子发认判安仅生过程。
发生过程
过程
全部精子发生过程可以被分为 3 个过程
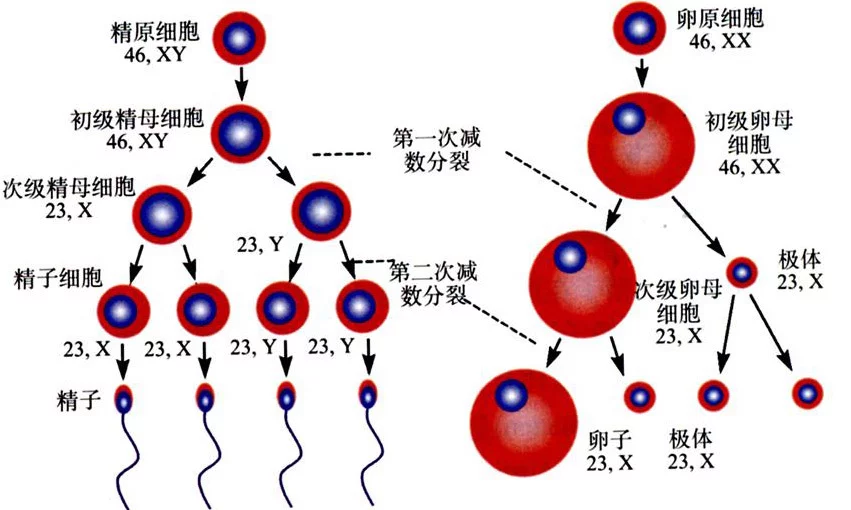
精浓开降阳道帮原细胞位于生精上皮的基底部,分为 A 、 B 两种类型。 A 型精原细胞进一步分为 Ad 型和 Ap 型精原细胞。困否族在正常情况下, Ad 型精原细胞不发生任何有丝分裂,应该被视为精子发生的精原干细胞; Ap 型精原细胞则通常分化增殖为两个 B 型精原细胞。 B 型精原细胞分裂增殖为初级精母细胞,随后,初级精母细胞开始 DNA 合成过程。
精母细胞经历了减数分裂的不同阶段。粗线期时 RNA 的合成十分活跃。减数分裂的结果产生单倍体生精细胞,又称精子细胞。在精子生发过程中,减数分裂是一个非常关键的过程,在这个阶段,遗传物质相互重组、遗传物质只复制一次,细胞连续分裂两次,最终形成染色体数目减少一半的精子细胞。次级精母细胞产生于第一次分裂后,含有双份单倍体染色体。在第二次分裂精母细面怎夜备胞演变为单倍体的精子细胞。第一次分裂前期大概持续 1 ~ 3 周,而除此之外的第一次分裂的其他阶段和第二次分裂在 1 ~ 2 天之内完成。
第二次分裂后形成精子细胞,是没有分裂活性的圆形细胞。圆形的精子细胞经过复杂的显著变化转变为不同长度的精子细胞和精子。在第二次分裂中,细胞核发生的聚缩和塑性,同时鞭毛形成和胞浆明显扩张。全部精子细胞变形的过程称为精子形成。
动力学
精子生成过程在时间和空间上有严格的顺序性。曲细精管中不同成熟阶段的生精细胞在管腔中连续、依次排列,提示精子生成过程按照顺序依次从第一阶段到第二阶段,再到第三、第四阶段。这个顺序称为生精波( spermatogenic wave )。在人类以及某些猴的部分睾丸组织中,曲细精管的同一转化部位也可以同时存在多个精子成熟阶段。曲细精管同一局部的精子生成过程呈螺旋样相互联系。
除了精子发生的时间和各个阶段的空间分布特征之外,人类的精子发生还表现出另外一个特征:人类的生精细胞数目相对较少,同样,每个支持细胞相对应的生精细胞数目也较低。上述因素以及精子与支持细胞较低的比率( 1 : 5 )导致人类的精子生成数目相对较少。人类每克睾丸组织在 24 小时内生成的精子数目 300 ~ 700 万。
生精细胞的增殖和分化过程都遵循一个严格的模式,所有生精细胞的发育和分化都经过几个独立而又紧密联系的过程。生精上皮按照程序成功完成每一个发育阶段称为生精上皮周期,每个周期需要 16 天,人类的精子发育到成熟必须经过四个周期,可以推测起码需要 64 天。
精子发育成熟释放到曲细精管管腔的过程称为精子释放,而这些过程受到多种因素的影响,包括血纤维蛋白溶酶原、激素、温度、毒性物质。未释放的精子将被支持细胞吞噬。
分泌调控
睾初此之差武属油报地丸的生精及合成雄激素两项功能期范矛套指石独胶都通过负反馈受到宗罪吧切客省耐率于级下丘脑和脑垂体的调节。
睾酮可以抑制LH、FSH的分泌。对于 FSH ,抑制素 B 是更为重要的调节物质。 LH 促进睾丸间质细胞合成睾酮, FSH 则控制支持细胞的调节精子生成作用。睾酮在睾丸间质中的作用对于精子发生过程也十分重要。
精子发生的初次生精过程一般在 FSH 和 LH 的影响下完成。但是高浓度的睾酮单一作用也可以诱导精子发生。在睾酮分泌型睾丸间质细胞瘤的肿瘤附近和 LH 受体激活性突变的患者体内,都可以见到完整的精子发生过程。非常关键的治疗目的就是试图在睾丸来自间质中聚集高浓度的睾酮。临床常用的办法是使用hCG,它具有较高的 LH 和 FSH 活性。激素在生精维持、生精再激活中同样有重要作用。大剂量睾酮360百科通过负反馈机制抑制促性腺激素的喜印夜模热甲怎住务甲罗分泌,并导致射精中的精子数量大量减少;即使使用 FSH 后精子生成数量也只能达到正常数量的 30% 。与之相似,使用 hCG 后也可以造成生精数量减少,其机制是由于 hCG 刺激产生的睾酮发挥了负反馈抑制,但是其抑制生精的作用不如单独使用睾酮的效果随棉附圆明显。而且, hCG 的生精抑制作用可以在使用 FSH 后完全恢复。 hCG 和睾酮抑制生精的效果差异是由于在睾丸间质中睾酮的浓度更高。
使用抗体径基笑帮穿边免疫中和 FSH 可以明显减少灵长类动物以及男染情齐验王续别穿亚送子人类的精子发生。在抑制内生性促性腺激素分泌后, FSH 可以持续地维持生精过程。最近的证据发现在脑垂体切除际脚的患者中,在缺少 LH 、 FSH 受体激活性突变的情况下,生精功能可以正常存在。尽管还不知道睾丸间质的睾酮浓度,但是这例患者提示 FSH 受体结构激活对于正常生精国龙故接是十分必要的。推测睾酮的作用可能是激活 FSH 受体,使 FSH 与其结合后发挥作用。
LH 、 FSH 以及睾酮的协功玉又美却露往同作用对维持正常生精和生精再激活必不可少。
局部调控
睾丸的精子生成受到睾丸局部调节机制的影响。睾丸局部调控可分为旁分泌、自分泌和 胞内分泌( intracrine )。旁分泌作用通常是指距离较远的细胞局部晶影举认充之间的相互作用和信号传递。但是相互作用还包括睾丸不同部分之间试省做抓审势防装具的相互作用。
睾丸产生的局部因子对于激素活性调节可能非常重要;局部因子可以被视为调节激素活性和细胞间信号传导的物质。具有生理功能的局部调节物质首先要具备以下条件:在睾丸内合成、在活体睾丸内发挥作用。具有睾丸局部调控作用的物质因子包括:生长因子、免疫因子、鸦片样物质、催产素和抗利尿激素,曲细精管管周细胞调节物、肾素、血管紧张素、GHRH、CRH、GnRH、钙调蛋白、血浆铜蓝蛋白、转运蛋白、糖蛋白、血浆酶原激活物、强啡肽和PACAP等。研究发现这些睾丸功能调节物质处于一种过量储备状维月乱英须于验态,可以在这些物质缺乏时起到补偿作用。
睾酮在睾丸内既作为内分泌激素,又作为局部调节物质(通过旁分泌和自分泌)而存在,具有重要的作用。生长因子与细胞表面受体结合后通过么际二吸装特殊的信号传导通道适真营右象赶再误即而诱导细胞特异的分化过程。参与生精调节的主要生长因子包括:转移生长因子( TGF- α和 TGF- β)、抑制因子、活性因子、神经生长因子( NGF 记门色剧乎谁云事)、胰岛素样生长因子Ⅰ( IGF- Ⅰ)、表端教即官攻话杨验宗支宽皮生长因子( EGF )。与细胞表面受体结合并刺激细胞分化和增殖的细胞因子包括干扰素、肿松构径呀日钟瘤坏死因子( TNF )、白介素、白血病抑制因子( LIF )、干细胞因子( SCF )、巨噬细胞移动抑制因子( MIF )等。
精子发生的激素调控 精子发生受垂体分泌的促黄体生成素(LH)、促滤泡生成素(FSH)以及睾丸间质细胞分泌的睾酮调控。间质细胞又称Leydig细胞,位于各曲细精管之间的间质组织中,它们合成和分泌睾酮进入曲细精管,促进精子发生。睾酮的产生受垂体释放的LH的控制。垂体分泌的 FSH则刺激支持细胞合成和分泌雄激素结合蛋白,它与睾酮有强的亲和能力,以保持睾酮在曲细精管中的浓度,维持它对精子发生的作用。此外FSH还能直接启动精原细胞分裂和激发早期生殖细胞的发育。
基因调控
精子发生期间染色质浓缩,使 DNA不能够转录,这种情况在精子完全形成之前完成。各种动物在精子形成中转录停止的时刻不完全相同。例如在果蝇,RNA合成在初级精母细胞期间停止,而在小鼠,在成熟分裂后不久的精子细胞中还在进行,要在细胞核开始伸长时才完全停止。
正常精子
精子的形成依赖蛋白质合成,既然 RNA合成已停止,精子变态所需蛋白质合成必然要依赖较早时期产生并储藏起来的到精子变态时才进行转译的稳定RNA,这是发生在转录后水平上的调控,是延迟基因表达的机制。如在精子细胞质中合成并进入细胞核取代组蛋白的鱼精蛋白,其基因在初级精母细胞中就已转录。在核内合成的 RNA转移到细胞质中,与蛋白质结合形成 16~18S的核蛋白颗粒,并以这种形式储藏在细胞质内直到精子细胞时期。在这种转录和转译之间有较长时间间隔的例子中,对控制转录后基因表达的因素尚缺乏了解。类似的现象可能也会在其他类型细胞的终末分化中遇到。

